 |

Гринченко С.Н.
Институт проблем информатики РАН, Москва
Within the framework of cybernetic biology problematics the new approach to a problem of alive systems adaptive behaviour (adaptation and evolution) modeling is offered on the basis of functioning of the hierarchical optimizational mechanism, is permanent tracing extremums of power character criterion functions on algorithms of adaptive random search.
Ключевые слова: иерархическая оптимизация, поисковая оптимизация, адаптация, Биосфера, биогеоценозы, организмы, органы, ткани, клетки, иерархические системы, моделирование, адаптивный случайный поиск.
Key words: hierarchical optimization, search optimization, adaptation, Biosphere, biogeocenosis, organism, organ, tissue, cell, hierarchical systems, simulation, adaptive random search.
В своих работах, посвященных кибернетическому подходу к развитию теоретической биологии, А.А.Ляпунов неоднократно указывал на важность проблемы иерархической организации живой природы (см., напр., [11]). Так, в работе "О кибернетических вопросах биологии" - доклада, доложенного на конференции по кибернетике еще в июне 1969 года в Новосибирске - он указывает: "Далеко идущая иерархичность организации управления в живой природе, а также ...иерархия ярусов организации живой природы определяют необходимость системного подхода к изучению живой природы. Фактически этот системный подход давным-давно осуществляется биологией. Однако логическая сторона дела далеко не достаточно выяснена. В связи с этим не выявлена возможная общность постановок вопросов в разных областях биологии, а также возможная общность подхода к различным биологическим явлениям и в особенности общность теоретических построений и математических моделей, которые могут быть использованы при изучении весьма разнообразных процессов жизнедеятельности" ([там же], стр.239).
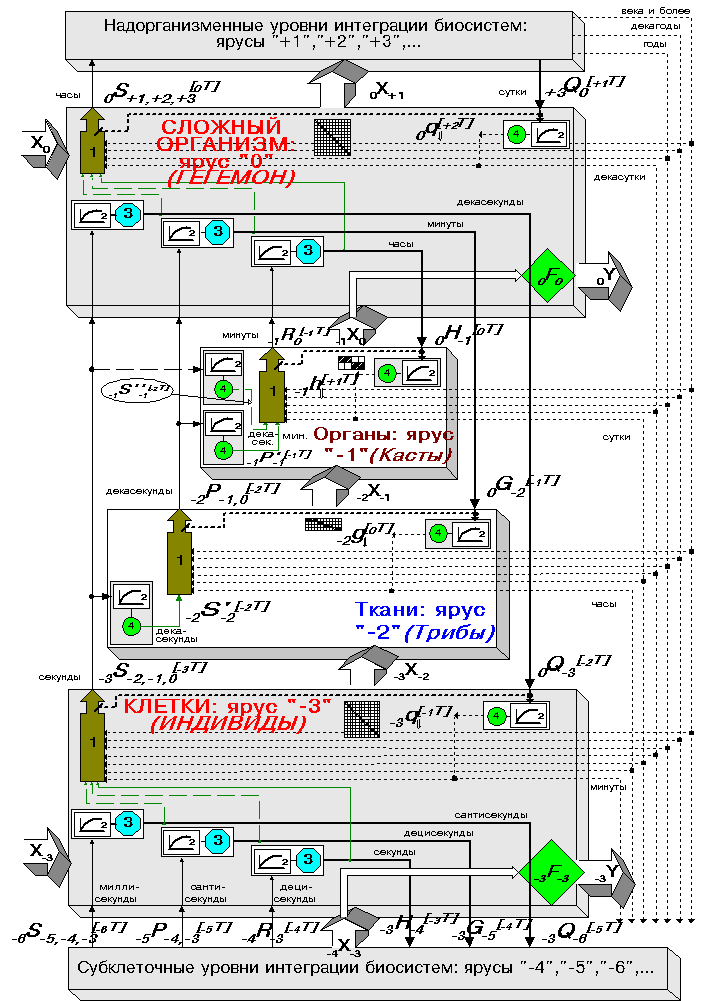
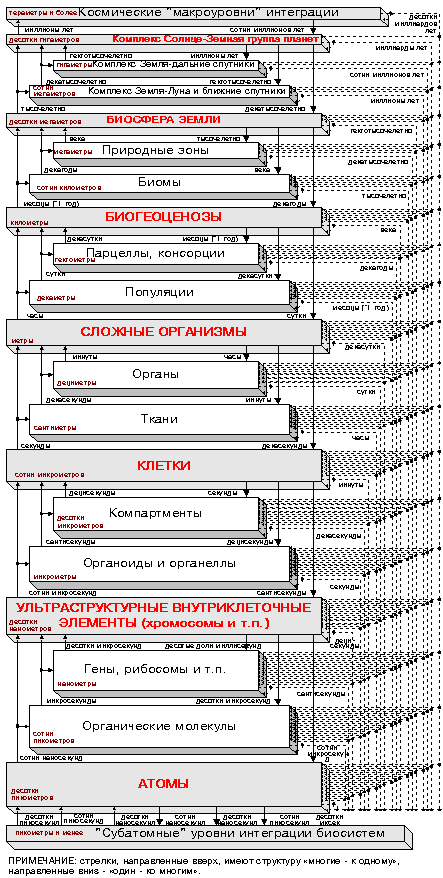
К сожалению, долгое время не уделялось достаточного внимания направлениям исследований, намеченным А.А.Ляпуновым в указанной, весьма актуальной, области знания. Тем не менее, интерес к подобным разработкам сохраняется. В частности, он инициировал проведение нами (с 1978-1979 года) ряда работ, в основе которых лежит предложение рассматривать модель механизмов реализации приспособительного поведения в биологических системах (т.е. их адаптации и эволюции) как иерархического механизма адаптивного случайного поиска, перманентно отслеживающего экстремумы целевых функций (энергетического характера), заданных на "основных уровнях биологической интеграции" - парадигма адаптивности (см., напр., [4-6] и библиографию к ним). Помимо "основных", в таком механизме выделяются пары "промежуточных уровней биологической интеграции", расположенные в иерархии между "основными" (см. рис. 1 и 2):
...; +9. Ближайший Космос (Солнце + земная группа планет); +8. Комплексы "Земля - дальние спутники и другие околоземные объекты"; +7. Комплексы "Земля - ближние спутники (включая Луну)"; +6. Биосфера Земли (экзосферы и магнитосферы планет); +5. Природные зоны; +4. Биомы; +3. Биогеоценозы; +2. Парцеллы, консорции; +1. Популяции; 0. Сложные организмы; -1. Органы; -2. Ткани; -3. Клетки; -4. Компартменты; -5. Органоиды, органеллы; -6.Ультраструктурные внутриклеточные элементы (хромосомы, вирусы и т.п.); -7. Макромолекулярные структуры (гены, рибосомы и т.п.); -8. Органические молекулы; -9. Атомы; ...
С нашей точки зрения, именно деятельность такого механизма является причиной возникновения значительного числа эмпирически наблюдаемых биологических ритмов.
Применительно к этой иерархической схеме ("супра-системе") задача
адаптации в ее отдельном иерархическом ("супра-") контуре (для примера -
это будет наиболее "наглядный" для человека супра-контур
"Сложный Организм-Клетки") иерархической природной системы может быть
записана (по аналогии с классической задачей адаптации сложных систем,
сформулированной Л.А. Растригиным [16]) следующим образом (см. рис. 2):
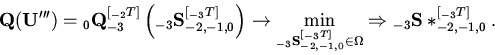
При этом:
![\begin{displaymath}
\Omega :{\left\{ {{\begin{array}{*{20}c}
{{\rm {\bf G}}({\r...
..._{0} ^{[{}_{ - 1}T]}) \ge 0} \hfill \\
\end{array}}} \right.}
\end{displaymath}](img4.gif)
Прежде чем перейти к подробным объяснениям обозначений, а также смысла и содержательных интерпретаций указанных рекуррентных соотношений, отметим главную особенность предлагаемой задачи - ее принципиальную трехконтурность. Каждый из трех целевых критериев (экстремальный, типа равенств и типа неравенств) замыкается на свою переменную, переменные же находятся в иерархической связи между собой (реализующие их ярусы "вложены" один в другой) - см. рис. 2.
Сквозная нумерация уровней, или ярусов, в иерархии природных систем широко используется нами при формальном описании задачи адаптации следующим образом:
Кроме того, каждой переменной мы приписываем указание на характерный темп ее
изменения. Эта информация "закодирована" как конструкция в виде верхнего
индекса с буквой T в квадратных скобках [
![]() T], расположенная справа от наименования переменной. Здесь вместо
точки - нижнего левого индекса при символе времени T -
подставляется номер темпа времени (в нашей классификации), характерный для
конкретного процесса: поисковой активности яруса в иерархии с соответствующим номером. Такое обозначение позволяет
наглядно видеть единый масштаб времени для сравнения темпов процессов
иерархической оптимизации самого различного характера - не только
поисковых, но и процессов изменения целевых критериев, а также темпов их
запоминания.
T], расположенная справа от наименования переменной. Здесь вместо
точки - нижнего левого индекса при символе времени T -
подставляется номер темпа времени (в нашей классификации), характерный для
конкретного процесса: поисковой активности яруса в иерархии с соответствующим номером. Такое обозначение позволяет
наглядно видеть единый масштаб времени для сравнения темпов процессов
иерархической оптимизации самого различного характера - не только
поисковых, но и процессов изменения целевых критериев, а также темпов их
запоминания.
Терминами ГЕГЕМОН и ИНДИВИДЫ (см. рис. 2) мы обозначаем те относящиеся к "основным уровням интеграции" ярусы в данном супра-контуре, первый из который задает целевые критерии для всех трех нижележащих ярусов, а второй - осуществляет активное поисковое поведение в субконтуре экстремального критерия, задающего возможность выбора (энергетически) наилучшего состояния системы из множества приемлемых. Терминами Трибы и Касты мы обозначаем те относящиеся к "промежуточным уровням интеграции" ярусы в данном супра-контуре, первый из который (с относительно однородной структурой) осуществляет поисковое поведение в субконтуре критерия типа равенств, диктующего системе (в соответствующих условиях) вполне определенное поведение - т.е. фактически законы природы, а второй (с относительно неоднородной структурой) - в субконтуре критерия типа неравенств, ограничивающего область приемлемых состояний системы.
Таким образом, символы
![]() здесь обозначают поисковые переменные процесса
иерархической оптимизации уровней клеточного, тканевого и органного
соответственно (т.е. ИНДИВИДОВ, Триб и Каст). Символы
здесь обозначают поисковые переменные процесса
иерархической оптимизации уровней клеточного, тканевого и органного
соответственно (т.е. ИНДИВИДОВ, Триб и Каст). Символы
![]() - это целевые критерии: соответственно экстремальный, типа
равенств и типа неравенств, задаваемые на организменном уровне (т.е.
ГЕГЕМОНА данного супра-контура). Символы
- это целевые критерии: соответственно экстремальный, типа
равенств и типа неравенств, задаваемые на организменном уровне (т.е.
ГЕГЕМОНА данного супра-контура). Символы
![]() - аналогичные целевые критерии, но задаваемые на
клеточном уровне (т.е. уровня ГЕГЕМОНА низшего супра-контура в иерархической
системе). Символы
- аналогичные целевые критерии, но задаваемые на
клеточном уровне (т.е. уровня ГЕГЕМОНА низшего супра-контура в иерархической
системе). Символы
![]() ,
,
![]() ,
,
![]() ,
,
![]() -
это память о траектории поиска и значениях поисковых параметров адаптивной
иерархической оптимизации, задаваемая на уровнях клеточном, организменном,
биогеоценотическом, биосферном и "ближайшего Космоса" соответственно (т.е.
уровнях ИНДИВИДОВ рассматриваемого и всех вышележащих супра-контуров в иерархической
системе), символы
-
это память о траектории поиска и значениях поисковых параметров адаптивной
иерархической оптимизации, задаваемая на уровнях клеточном, организменном,
биогеоценотическом, биосферном и "ближайшего Космоса" соответственно (т.е.
уровнях ИНДИВИДОВ рассматриваемого и всех вышележащих супра-контуров в иерархической
системе), символы
![]() ,
,
![]() - память процесса адаптивной иерархической оптимизации, задаваемая на
уровнях тканевом, популяционном, биомном и "комплекса Земля-Луна"
соответственно (т.е. уровнях Триб рассматриваемого и всех вышележащих
супра-контуров в иерархической системе), символы
- память процесса адаптивной иерархической оптимизации, задаваемая на
уровнях тканевом, популяционном, биомном и "комплекса Земля-Луна"
соответственно (т.е. уровнях Триб рассматриваемого и всех вышележащих
супра-контуров в иерархической системе), символы
![]() - память процесса адаптивной
иерархической оптимизации, задаваемая на уровнях органном,
парцелло-консорционном, природных зон и "комплекса Земля-дальние спутники"
соответственно (т.е. уровнях Каст рассматриваемого и всех вышележащих
супра-контуров в иерархической системе). Наконец, символы
- память процесса адаптивной
иерархической оптимизации, задаваемая на уровнях органном,
парцелло-консорционном, природных зон и "комплекса Земля-дальние спутники"
соответственно (т.е. уровнях Каст рассматриваемого и всех вышележащих
супра-контуров в иерархической системе). Наконец, символы
![]() отражают формы
внутреннего восприятия и преобразования поисковых процессов на органном и
тканевом уровнях соответственно (т.е. уровнях Каст и Триб данного супра-контура).
Ограниченность объема настоящей статьи не позволяет привести здесь подробную
интерпретацию всех вышеуказанных переменных в терминах наук, традиционно их
изучающих.
отражают формы
внутреннего восприятия и преобразования поисковых процессов на органном и
тканевом уровнях соответственно (т.е. уровнях Каст и Триб данного супра-контура).
Ограниченность объема настоящей статьи не позволяет привести здесь подробную
интерпретацию всех вышеуказанных переменных в терминах наук, традиционно их
изучающих.
Предложение использовать целевой подход в предлагаемой трактовке для описания процессов приспособительного поведения в биосистемах опять-таки коррелирует с позицией А.А. Ляпунова, который, обсуждая вопрос об изучении информационных потоков, управляющих эволюцией управляющих систем, формулирует: "Эти задачи обладают тем свойством, что цель управления возникает внутри системы, а не задается извне. В основе лежит то обстоятельство, что случайно возникающие приспособления, содействующие самосохранению, ведут к повышенной устойчивости объекта и закрепляются путем отбора" ([11], стр. 216). Правда, есть и существенные отличия: мы считаем, что, "путем отбора закрепляются" не просто "приспособления, содействующие самосохранению", но приспособления, соответствующие при этом тенденции движения к экстремуму энергетического критерия соответствующих уровней (ярусов) в иерархии. Для систем клеточного уровня эта идея предложена нами и наиболее полно изложена в [2-3], для систем организменного уровня ее развивает В.Д. Цветков [18], для систем надорганизменного уровня подобная идея предложена и обоснована Н.С. Печуркиным [12].
В качестве алгоритма иерархической оптимизации наиболее адекватной моделью является случайный поиск в терминологии Л.А. Растригина [14-15]. Его теория является важным элементом раздела технической кибернетики - теории поисковой оптимизации (экстремального регулирования), которую в 1960-1970-х гг. создал ряд отечественных и зарубежных ученых, среди которых можно выделить Д.И. Батищева, Ю.М. Ермольева, В.В. Казакевича, А.А. Красовского, Ю.И. Неймарка, А.А. Первозванского, Л.А. Растригина, Р.Г. Стронгина, А.А. Фельдбаума, Я.З. Цыпкина, У.Р. Эшби, Д.Б. Юдина. В простейших случаях случайный поиск генерирует в некоторой окрестности поисковой переменной ее случайное приращение, вычисляет значение оптимизируемой функции, сравнивает с ее значением на предыдущем шаге поиска и, в зависимости от результата (приближения/удаления от экстремума), либо повторяет предыдущий шаг, либо генерирует новое случайное приращение поисковой переменной; далее все повторяется. В этих случаях его, безусловно, можно рассматривать как реализации вариантов простейшего оптимизационного метода "проб и ошибок".
В более сложных случаях этот алгоритм обладает свойством адаптивности, реализуемом с помощью запоминания как траектории поискового процесса (значений поисковых переменных и целевых функций, а также соответствующих ограничений), так и параметров самого алгоритма (величины и направления поискового шага, глубины памяти и т.п.). Именно это свойство делает его весьма эффективным средством решения сложных экстремальных задач самого различного типа. Более того, именно механизм адаптивного случайного поиска является средством - причем, по нашему мнению, единственно адекватным - моделирования поведения сложных природных систем. Действительно, довольно трудно представить себе некоторый живой объект, адаптивное поведение которого (например, значение градиента целевой функции) рассчитывалось бы в каком-то гипотетическом вычислительном блоке (что необходимо для любого регулярного метода оптимизации). Применительно же к адаптивному случайному поиску - это вполне реально, поскольку он не требует никаких подобных вычислений, а оценивает искомый градиент на базе нескольких имеющихся конкретных реализаций целевой функции. Добавим для читателя, знакомого с теорией автоматов, что алгоритм случайного поиска легко формулируется на языке вероятностных автоматов - это просто два разных языка описания одной и той же сущности.
Отметим, что в рамках предлагаемой концепции случайность мы рассматриваем как средство отражения
наблюдаемой в реальности иерархичности природных систем. Последнее можно трактовать как
обобщение достаточно часто используемой на практике гносеологической
ипостаси случайности как меры нашего незнания об объекте. Различие состоит в том, что в иерархической
оптимизационной подсистеме некоторого уровня интеграции А даже при вполне
регулярном поведении каждой из составляющих ее подсистем a![]() ,...,a
,...,a![]() (обладающих свойством активности) рассмотрение
такого поведения "в целом" - одновременно и всех сразу, т.е. "с точки
зрения" подсистемы именно высшего уровня А, - может выглядеть только как
случайное: у подсистемы А просто нет так называемого "планшета" для
фиксации поведения всех ее подсистем a
(обладающих свойством активности) рассмотрение
такого поведения "в целом" - одновременно и всех сразу, т.е. "с точки
зрения" подсистемы именно высшего уровня А, - может выглядеть только как
случайное: у подсистемы А просто нет так называемого "планшета" для
фиксации поведения всех ее подсистем a![]() ,...,a
,...,a![]() .
.
В частности, для супра-контура "Сложный организм-клетки" (поскольку мы относимся именно к организменному уровню) таковая мера (нашего) незнания сложного организма о поведении всех клеток, из которых он состоит, объективна и абсолютна, а в обыденном варианте (частном случае по отношению к предлагаемому, т.е. "организм-организм") она субъективна (мера НАШЕГО незнания о каком-либо явлении НАШЕГО уровня интеграции) и относительна [6]. Нам представляется, что целесообразно говорить о выявлении в гносеологическом аспекте интерпретации понятия случайности некоторого конкретного варианта (направления), который можно было бы назвать "релятивно-иерархическим".
Итак, нами введен некий формализм, опирающийся на представление процессов в биологической системе - в частности, в организме - как итеративного оптимизационного процесса, происходящего сразу в нескольких характерных темпах. Эти темпы определяются "привязкой" соответствующих процессов как к тому или иному уровню в иерархии природной системы, так и к его месту в оптимизационном супра-контуре (рис. 1-2).
Весьма важное значение имеет тот факт, что значения этих характерных времен, или темпов, супра-оптимизационных процессов сводятся (в среднем) к равномерной последовательности, с шагом примерно на порядок. Другими словами, мы постулируем, что "в модели иерархической оптимизации природных систем (реализации их приспособительного поведения) механизмы восприятия и интеграции специфической информации, а также механизмы выработки памяти характеристик оптимизационных процессов, связывающих смежные уровни интеграции (ярусы) в системе, обладают свойством инерционности ("сглаживания" темпа изменения) на некоторую величину, примерно равную 10-20". Последнее значение соответствует практике применения адаптивных алгоритмов поисковой оптимизации в технике (в частности, при решении задач автоматизированного проектирования различных технических конструкций). Эта практика показывает, что максимальная эффективность оптимизационного процесса достигается, если характерное замедление двух смежных процессов адаптации (первичной, или собственно оптимизации - параметрической - структурной) составляет примерно 10-20 раз.
В свое время А.В. Жирмунский и В.И. Кузьмин, изучая критические режимы в
биосистемах, собрали огромное число экспериментальных фактов,
свидетельствующих о том, что темпы большого числа биологических процессов
соотносятся как e![]() = 15,15426... [7]. Более того, им удалось
получить эту величину аналитически (не привлекая, впрочем, для этого
каких-либо предположений об оптимизационном характере соответствующих
процессов).
= 15,15426... [7]. Более того, им удалось
получить эту величину аналитически (не привлекая, впрочем, для этого
каких-либо предположений об оптимизационном характере соответствующих
процессов).
Опираясь на тот факт, что рассматриваемые нами супра-оптимизационные
процессы в природных системах также могут рассматриваться как содержащие
некие критические точки, а также на близость указанной величины к интуитивно
необходимому и практически подобранному (для "развязки" управленческих
процессов во времени) изменению "на порядок", мы и интерпретируем величину соотношения темпов
- относительно дискретных характерных времен, или временных шагов, с которыми изменяются соответствующие
оптимизационные процессы в иерархической системе - как величину, близкую к
e![]() (это же справедливо и для пространственных характеристик
природных систем - см. рис. 1).
(это же справедливо и для пространственных характеристик
природных систем - см. рис. 1).
Анализ концепции иерархической оптимизации как модели универсального механизма реализации приспособительного поведения в живой природе позволяет наметить некоторые ее следствия, а именно:
Рассмотрим, каково положение предлагаемой концепции в системе общенаучного знания. В идейном плане она может рассматриваться как одна из реализаций достаточно хорошо разработанного кибернетического направления, связанного с телеологическими/телеономическими представлениями о поведении сложных систем. Это демонстрирует, в частности, ряд публикаций Н.Н. Моисеева.
Это перекликается и с недавно появившимися в философской литературе предложением (восходящим к Анаксагору и Платону и базирующимся на огромном материале последующих поколений исследователей всех отраслей знания) о конституировании новой трансдисциплинарной науки - оптимологии [13]. Автор этого предложения О.С. Разумовский отмечает, что "исходной предпосылкой оптимологии должен быть эмпирический факт: главный вектор существования, организации, функционирования и развития всех живых систем, общества и человека, а также смешанных или гибридных систем (...) - интенция к оптимизации (к оптимуму)" [там же]. При этом в качестве одной из самых трудных методологических проблем он рассматривает проблему выбора прототипа для идеального объекта оптимологии, без решения которой нельзя "вообще конструировать понятийно-номологический базис теории" [там же]. Имеется в виду выбор единого прототипа, к которому можно было бы потом редуцировать все остальное типы бихевиоральных (поведенческих) систем. Позволим себе высказать предположение, что в качестве базиса выбора такого прототипа (фактически, языка, адекватного оптимизационному подходу или оптимизационному мышлению) как раз и может выступить предлагаемая концепция [4].
В свою очередь, анализируя предмет биологической кибернетики, А.А. Крушанов констатирует, что последний "состоит в изучении специфических для живых существ общих принципов и конкретных механизмов целесообразного саморегулирования и активного взаимодействия с окружающей средой. Следует подчеркнуть, что при этом речь идет о разделе именно биологии, о более глубоком изучении явлений жизни, о вскрытии новых ее закономерностей средствами кибернетического анализа, а не об иллюстрации общих законов кибернетики на примерах деятельности живых существ. Поэтому, строго говоря, было бы правильнее определить эту область знания как кибернетическую биологию ..." [9]. По-видимому, данная точка зрения может быть с успехом отнесена и к предлагаемой разработке.
Итак, с позиций предлагаемой кибернетической концепции "иерархической оптимизации" важнейшее свойство биологических систем - их иерархичность - рассматривается нами как средство, необходимое для реализации фундаментального свойства природных систем - их адаптивности. Именно и только иерархичность пространственно-временных характеристик позволяет "разнести" в пространстве и во времени весь спектр процессов адаптации природных систем.
Таким образом, справедливость положения А.А. Ляпунова о том, что "... кибернетический подход к изучению процессов жизнедеятельности в известном смысле дополняет описательно-биологический и молекулярно-биохимический подход, т.к. точное описание процессов управления, и только оно, позволяет судить о том, достаточно ли полно выявлены элементарные физико-химические жизненные акты для объяснения целостной картины, наблюдаемой в биологии" (см. [11], стр. 220) получает новое подтверждение.
 Ваши комментарии
Ваши комментарии
|
![[SBRAS]](/gif/s_sigma.gif)
[Головная страница] [Конференции] [СО РАН] |
© 2001, Сибирское отделение Российской академии наук, Новосибирск
© 2001, Объединенный институт информатики СО РАН, Новосибирск
© 2001, Институт вычислительных технологий СО РАН, Новосибирск
© 2001, Институт систем информатики СО РАН, Новосибирск
© 2001, Институт математики СО РАН, Новосибирск
© 2001, Институт цитологии и генетики СО РАН, Новосибирск
© 2001, Институт вычислительной математики и математической геофизики СО РАН, Новосибирск
© 2001, Новосибирский государственный университет
Дата последней модификации Wednesday, 05-Sep-2001 15:36:11 NOVST