Евстафьев В.К.
Лимнологический институт СО РАН
Формализация описания динамики популяций в природной среде, несомненно, одна
из важнейших задач современного естествознания. Экологические проблемы,
вставшие перед человечеством, заставляют внимательнее и вдумчивее относиться
к природным процессам. Появилась настоятельная необходимость в моделях,
описывающих реальные экосистемы и обладающих надежной предсказательной
силой. Здесь возможны (и реализуются) два подхода к решению задачи как в
случае описания газа в сосуде, где в принципе можно было бы составить
![]() уравнений движения для каждой молекулы и, таким
образом, зная их координаты и скорость в любой момент времени, знать
состояние газа в целом. Но решение столь фантастически громоздкой системы
уравнений, разумеется, практически невозможно осуществить. Посему
реализовался другой подход - термодинамический (кибернетический), где нас
интересуют не внутренние процессы, а только макропараметры системы. Для газа
это давление
уравнений движения для каждой молекулы и, таким
образом, зная их координаты и скорость в любой момент времени, знать
состояние газа в целом. Но решение столь фантастически громоздкой системы
уравнений, разумеется, практически невозможно осуществить. Посему
реализовался другой подход - термодинамический (кибернетический), где нас
интересуют не внутренние процессы, а только макропараметры системы. Для газа
это давление ![]() , объем
, объем ![]() , температура
, температура ![]() , связанные известным уравнением
Менделеева-Клайперона
, связанные известным уравнением
Менделеева-Клайперона
![]() - газовая постоянная.
- газовая постоянная.
Примерно такая же ситуация складывается сейчас в экологии. Модели на основе простых представлений не могут удовлетворить требований адекватности описания экосистем. Поэтому развитие этих моделей пошло по пути все большей детализации. Это несколько улучшает их точность, но при этом они становятся математически столь громоздкими, что с ними уже трудно работать. Этим, думается, объясняется та пропасть, что разверзлась между модельерами и биологами-практиками.
В докладе мне хотелось бы обсудить старый термодинамический метод применительно к описанию экосистем.
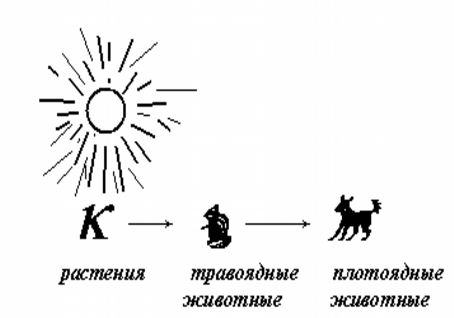
Здесь и далее будем считать, что в математически формализованном виде популяции представляют собой множество однородных самовоспроизводящихся биологических особей (в случае необходимости вводятся половое различие, дифференциация по возрасту и т.д.). Источником энергии для растительных популяций является солнечное излучение, для животных - энергия пищи, т.е. других съеденных особей. Цепочку популяций, связанных пищевыми отношениями, принято называть трофической цепью. Такие линейные цепи встречаются в энергетически бедных изолированных экосистемах, например, полярных, но в большей части биосферы они опутаны множеством боковых ответвлений, поэтому правильнее говорить уже о трофических сетях, а в конечном счете о единой биосферной пищевой сети (рис. 1).
С точки зрения физики трофическая цепь интересна тем, что представляет собой однонаправленный канал проводимости. Вследствие сезонной неоднородности поступления энергии в трофическую цепь через первичное фотосинтезирующее звено, вдоль этой трофической цепи генерируются волны изменения численности популяций, бегущие от начального к верхнему звену. Т.е. к описанию такой системы вполне применим аппарат волновой механики.
Вследствие вращения Земли вокруг собственной оси и вокруг Солнца мы наблюдаем суточную и годовую периодичность поступления солнечного света и тепла к поверхности нашей планеты. Эта периодичность обусловливает и биологическую суточную и годовую ритмику. Весенне-летняя вегетация растений создает обилие пищи для травоядных животных и, соответственно, ведет к увеличению их численности. Они в свою очередь служат пищей для плотоядных хищников. В результате импульс энергии, фотосинтетически аккумулированной растениями, передается по пищевой цепи в виде бегущей волны всплеска численности популяций.
Возникает вопрос: какова дальнейшая судьба бегущих волн, достигших последнего трофического звена? Возможны два варианта: либо волна бесследно диссипирует, либо возникает какая-то обратная связь с первичным растительным звеном, вследствие чего ее путь замыкается в кольцо. Заманчиво предположить второй сценарий с построением красивой физической модели круговой циклической динамики, подобно атомной, со всеми вытекающими резонансными последствиями. Однако в отношении годовой периодичности такая обратная связь не несет никакой информационной роли из-за строгого постоянства длины года и, соответственно, если даже она была на заре эволюции, то к настоящему времени должна была бы совершенно рудиментироваться. Т.е. в отношении годовых циклов мы должны согласиться с выводом об отсутствии обратной связи между верхним и нижним звеньями трофической цепи.
А если какое-то важное для растений периодическое воздействие не строго постоянно? Например, циклы солнечной активности. За время с 1705 по 2000 годы средняя длина циклов составила 10.93 года, варьируя от 8 до 15 лет нормальным гауссовским образом. В этом случае роль обратной связи представляется значимой. Мы со старшим научным сотрудником Лимнологического института СО РАН Ниной Александровной Бондаренко проанализировали многолетнюю динамику основной байкальской водоросли Aulacoseira baicalensis за отрезок времени с 1946 по 1999 годы и нашли, что она резонансно сопряжена с периодом в 22 года, который есть период инверсии магнитных полюсов Солнца, более известный как 11-летние циклы солнечной активности. Физическая основа (резонансная) явления и результаты практического анализа опубликованы нами в журнале ''Биофизика'', 6, 2000, стр. 1089-1095, поэтому останавливаться на ее разъяснении не будем, а сразу перейдем к теме доклада.
В соответствии с нашей моделью, которая была основана на 11-летней повторяемости максимумов солнечной активности, очередная вспышка развития весеннего фитопланктона озера Байкал должна была произойти в 2001 году, но неожиданно она случилась в 2000-ом. В чем дело? Простая арифметика показывает, что средний цикл солнечной активности в 20 веке составил 10.5 лет. Но если с 1946 по 1990 годы динамика мелозиры протекала в ритме 11 лет, то где-то должна была проявиться и ритмика в 10 лет. К сожалению, количественных данных по байкальскому фитопланктону за первую половину столетия нет, что сильно затрудняет проверку гипотезы. Есть лишь некоторые косвенные сведения. М.М. Кожов, один из видных исследователей Байкала довоенного, военного и послевоенного времени, отмечает 1943 год как ''мелозирный'', который по нашей модели должен был бы быть в 1942 году. Это облачко, по меткому замечанию Лоренца, которому мы поначалу не придали серьезного значения, в 2000 году превратилось в грозовую тучу. Если несоответствие 1942-43 годов можно было отнести на счет какой-то ошибки, скрытой глубиной времени, то несоответствие 2000 и 2001 годов заставило нас уточнить модель с учетом того, что солнечная активность не строго 11-летняя, а способна варьировать по длине. Мы рассматривали три сценария:
- периодичность вспышек фитопланктона за длительное время эволюции адаптировалась к среднему ритму солнечной активности, т.е. 11 годам, и в настоящее время не меняется;
- адекватно меняется вслед за изменениями интервалов солнечной активности;
- фитопланктон сформировал основную частоту, адаптированную к среднему периоду солнечной активности 11 лет, и две запасные слева и справа, т.е. 10 и 12 лет.
Второй сценарий мы отбросили сразу как не экономичный и не согласующийся с
практикой. Первый, вследствие своей консервативности, также маловероятен.
Третий представляется нам наиболее вероятным. Т.к. средняя ритмичность
солнечной активности за 20 век составила 10.5 лет, то в динамике
фитопланктона должны были бы проявиться частоты 11 и 10 лет. 2000 год
выдался ''мелозирным'' (доминировали A. skvortzowii, Stephanodiscus binderanus, A. baicalensis), т.е. выскочил 10-летний цикл, в то
время как согласно 11-летней цикличности вспышка ожидалась весной 2001 года.
Наше объяснение наблюдаемому феномену заключается в том, что в 1990 узловом
году, когда происходила закладка следующего базового цикла, ''мелозирный
год'' пришелся на ниспадающую ветвь кривой пика солнечной активности
![]() , и фитопланктон отреагировал на укорочение солнечного
цикла активизацией своей запасной 10-летней частоты (табл.).
, и фитопланктон отреагировал на укорочение солнечного
цикла активизацией своей запасной 10-летней частоты (табл.).
| Узловые ''мелозирные годы'' | Солнечная активность
|
||
| 33 | 1945 | ||
| 1946 | 93 | 1946 | +60 |
| 151 | 1947 | ||
| 142 | 1956 | ||
| 1957 | 190 | 1957 | +48 |
| 185 | 1958 | ||
| 94 | 1967 | ||
| 1968 | 106 | 1968 | +12 |
| 106 | 1969 | ||
| 93 | 1978 | ||
| 1979 | 155 | 1979 | +62 |
| 155 | 1980 | ||
| 158 | 1989 | ||
| 1990 | 143 | 1990 | -15 |
| 146 | 1991 | ||
| 93 | 1999 | ||
| 2000 | 120 | 2000 | +27 |
| <120 | 2001 | ||
Будет следующий цикл 10-летним или фитопланктон вернется к своему основному
ритму в 11 лет - покажет время: в первом случае весенние вспышки будут
наблюдаться в 2004, 2007 и 2010 годах, во втором - в 2004, 2007 и 2011.
Исходя из того, что ![]() между 1999 и 2000 годами положителен, +27, то
следует ожидать возвращения фитопланктона к основной частоте. Т.е. смысл
уточнения к нашей модели заключается в том, что фитопланктон Байкала
функционирует на основной частоте в 11 лет, пока
между 1999 и 2000 годами положителен, +27, то
следует ожидать возвращения фитопланктона к основной частоте. Т.е. смысл
уточнения к нашей модели заключается в том, что фитопланктон Байкала
функционирует на основной частоте в 11 лет, пока ![]() положителен, и
укорачивает цикл до 10 лет, если
положителен, и
укорачивает цикл до 10 лет, если ![]() становится отрицательным. Таков,
как кажется логичным, механизм, посредством которого фитопланктон удерживает
в максимуме солнечной активности свои узловые генеративные годы.
становится отрицательным. Таков,
как кажется логичным, механизм, посредством которого фитопланктон удерживает
в максимуме солнечной активности свои узловые генеративные годы.
Таким образом, простые термодинамические соображения, примененные даже к столь сложным системам как экосистемы, могут приводить к важным результатам. На примере ''мелозирных лет'' байкальского фитопланктона, которые длительное время оставались загадкой для лимнологов, в докладе, надеюсь, убедительно было показано могущество этого старого метода.
 Ваши комментарии
Ваши комментарии
|
![[SBRAS]](/gif/s_sigma.gif)
[Головная страница] [Конференции] [СО РАН] |
© 2001, Сибирское отделение Российской академии наук, Новосибирск
© 2001, Объединенный институт информатики СО РАН, Новосибирск
© 2001, Институт вычислительных технологий СО РАН, Новосибирск
© 2001, Институт систем информатики СО РАН, Новосибирск
© 2001, Институт математики СО РАН, Новосибирск
© 2001, Институт цитологии и генетики СО РАН, Новосибирск
© 2001, Институт вычислительной математики и математической геофизики СО РАН, Новосибирск
© 2001, Новосибирский государственный университет
Дата последней модификации Thursday, 27-Sep-2001 18:19:13 NOVST