
Галимзянов А.В.
Институт биологии Уфимского научного центра РАН
К настоящему времени накапливаются большие массивы экспериментальных данных по генным сетям эукариот, управляющим различными процессами, в частности, онтогенетическими, что позволяет строить математические модели этих систем и с их помощью изучать особенности механизмов, лежащих в основе развития того или иного организма. Настоящий доклад посвящен изучению подсистемы управления морфогенезом цветкаArabidopsis thaliana (AT-ПУМ) и генной подсети, управляющей ранним онтогенезом Drosophila melanogaster (Dr-УГС), на основе метода обобщенных пороговых моделей (ОПМ) (Tchuraev, 1991; Чураев, 1993). Исследование моделей проводилось с помощью оригинального сопровождающего программного обеспечения "Аnalyzer of the Gene Network Dynamics" (AGENDY) (Galimzyanov, 2000), представляющего собой компьютерную среду для построения моделей эукариотических геных сетей произвольной сложности и расчета динамики в моделях с учетом значений кинетических параметров (единичных интенсивностей синтеза продуктов генов, коэффициентов деградации мРНК и белков, временных задержек, пороговых концентраций белков). При построении моделей AT-ПУМ-2 и Dr-УГС-1 правдоподобные границы значений кинетических параметров оценивались на основе известных экспериментальных данных (Патрушев, 2000; Ashburner, 1990; Driever, Nüsslein-Volhard, 1988; Manoukian A.S., Krause, 1992).
Качественное описание системы. Развитие цветка Arabidopsis состоит из двух последовательных процессов - перехода к
цветению и органогенеза (рис.1) (Levy, Dean, 1998, review; Haughn et al.,
1995, review). Переход к цветению Arabidopsis контролируется тремя основными
сигнальными путями: фотопериодическим, автономным и гиббереллин-зависимым
(Abel et al., 2000). Специфический баланс между внешними и внутренними
факторами через соответствующие сигнальные пути приводит к активации
транскрипции или подавлению активности генов, которые задают свойства
цветковой меристемы и управляют органогенезом. Цветок Arabidopsis состоит из четырех
концентрических колец (или мутовок): w![]() , w
, w![]() , w
, w![]() и w
и w![]() , расположенных вокруг оси стебля снаружи внутрь
(w
, расположенных вокруг оси стебля снаружи внутрь
(w![]() - внешнее, w
- внешнее, w![]() - внутреннее). В каждом кольце
формируются органы одного типа: четыре чашелистика - в первой мутовке,
четыре лепестка - во второй, шесть тычинок - в третьей и два плодолистика
- в четвертой.
- внутреннее). В каждом кольце
формируются органы одного типа: четыре чашелистика - в первой мутовке,
четыре лепестка - во второй, шесть тычинок - в третьей и два плодолистика
- в четвертой.
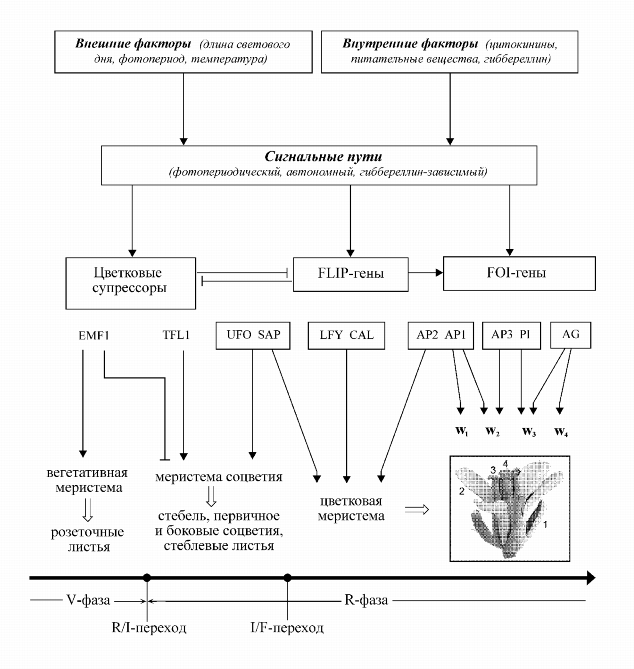 |
Индукция цветения переключает вегетативную фазу развития (V-фаза)
побега на репродуктивную (R-фаза) и приводит к образованию мериcтемы
соцветия (R/I-переход - от "rosette/inflorescence"). Далее апикальная
меристема репродуктивного зачатка преобразуется в цветковую меристему
(I/F-переход - от "inflorescence/flower"). Из цветковой меристемы в
течение 12 стадий формируются органы цветка. R/I и I/F-переходами управляет
контроллер фазового переключения (Haughn et al., 1995); продукты генов
класса FLIP задают свойства цветковой меристемы; гены класса FOI управляют
органогенезом. Позитивная и негативная регуляция активности элементов
системы обозначается стрелками ![]() и +. Изображение цветка взято из
банка данных Munich Information Сenter for Protein Sequences
(http://mips.gsf.de). 1 - чашелистик, 2 - лепесток, 3 - тычинка, 4 -
пестик.
и +. Изображение цветка взято из
банка данных Munich Information Сenter for Protein Sequences
(http://mips.gsf.de). 1 - чашелистик, 2 - лепесток, 3 - тычинка, 4 -
пестик.
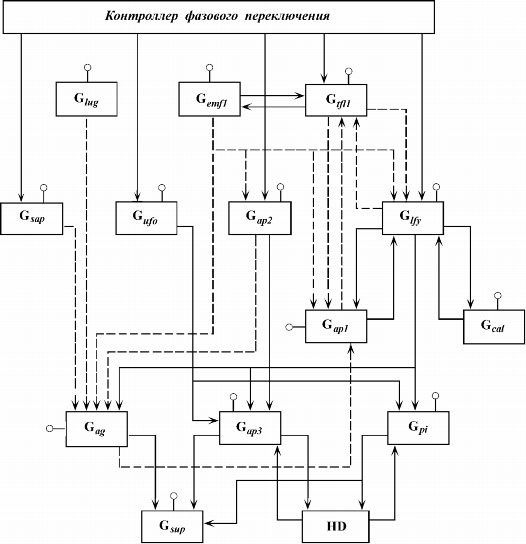
Построение и анализ модели. В модель AT-ПУМ-2 входят синтетические блоки G![]() , G
, G![]() ,
..., G
,
..., G
![]() - блоки синтеза белковых продуктов генов
EMF1, TFL1, LFY, AP1, AP2, CAL, UFO, LUG, SAP, AG, AP3, PI, SUP соответственно и блок HD формирования белок-белкового комплекса AP3/PI.
Каждый синтетический блок описывается как генетический блок в методе ОПМ
(Tchuraev, 1991; Чураев, 1993), а блок HD - с привлечением аппарата
химической кинетики. Булевы функции комбинаторов строятся в соответствии с
логикой регуляторных взаимодействий между белками и генами реальной системы.
Остальные элементы информационной микроструткуры генетического блка имеют
стандартный вид. Блок-схема модели изображена на рис. 2.
- блоки синтеза белковых продуктов генов
EMF1, TFL1, LFY, AP1, AP2, CAL, UFO, LUG, SAP, AG, AP3, PI, SUP соответственно и блок HD формирования белок-белкового комплекса AP3/PI.
Каждый синтетический блок описывается как генетический блок в методе ОПМ
(Tchuraev, 1991; Чураев, 1993), а блок HD - с привлечением аппарата
химической кинетики. Булевы функции комбинаторов строятся в соответствии с
логикой регуляторных взаимодействий между белками и генами реальной системы.
Остальные элементы информационной микроструткуры генетического блка имеют
стандартный вид. Блок-схема модели изображена на рис. 2.
 |
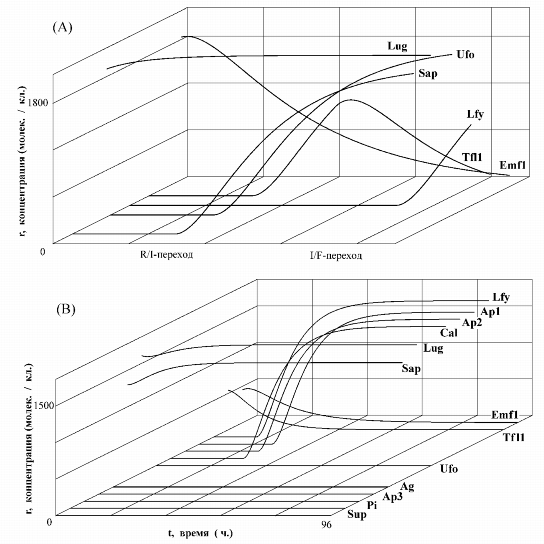
Изучение функционирования математической модели AT-ПУМ-2 проводилось с
помощью пакета программ AGENDY. Посредством компьютерных экспериментов были
выявлены один переходный режим R![]() (рис. 3,A), четыре
стационарных состояния (аналоги устойчивых точек типа "узел") R
(рис. 3,A), четыре
стационарных состояния (аналоги устойчивых точек типа "узел") R
![]() (рис. 3,B), R
(рис. 3,B), R
![]() , R
, R
![]() и R
и R
![]() и один неустойчивый синхронный режим,
впервые найденный ранее (Чураев, Ратнер, 1973).
и один неустойчивый синхронный режим,
впервые найденный ранее (Чураев, Ратнер, 1973).
 |
Стационарные режимы R
![]() , R
, R
![]() ,
R
,
R
![]() , R
, R
![]() модели АТ-ПУМ-2 легко
интерпретируются: согласно генетической ABC-модели (Coen, Meyerowitz, 1991),
режим R
модели АТ-ПУМ-2 легко
интерпретируются: согласно генетической ABC-модели (Coen, Meyerowitz, 1991),
режим R
![]() приводит к формированию чашелистиков
(w
приводит к формированию чашелистиков
(w![]() , R
, R
![]() - лепестков (w
- лепестков (w![]() ,
R
,
R
![]() - тычинок (w
- тычинок (w![]() , R
, R
![]() - плодолистиков (w
- плодолистиков (w![]() . Переходный режим R
. Переходный режим R![]() в
модели соответствует стадии формирования меристемы соцветия в промежуток
времени между R/I- и I/F-переходами. Синхронный режим может возникать на
границе между областями w
в
модели соответствует стадии формирования меристемы соцветия в промежуток
времени между R/I- и I/F-переходами. Синхронный режим может возникать на
границе между областями w![]() и w
и w![]() во время
инициации примордиев лепестков и тычинок.
во время
инициации примордиев лепестков и тычинок.
Как отмечено в докладе (Чураев, 2001), адекватные модели реальных процессов
должны обладать параметрической устойчивостью. В целях тестирования
параметрической устойчивости модели AT-ПУМ-2 было проведено три вида
компьютерных экспериментов. В первом эксперименте значения параметров
случайно выбирались в интервалах (x![]() - 0,3x
- 0,3x![]() ,
x
,
x![]() + 0,3x
+ 0,3x![]() , где x
, где x![]() - параметр из набора
со значением, меньшим нижней границы соответствующего правдоподобного
интервала. Во втором эксперименте значения параметров случайно выбирались в
пределах 30% отклонения от "хорошего" набора параметров. В качестве
"хорошего" набора параметров выбран такой, в котором значения всех
переменных лежат в середине правдоподобных интервалов. В обоих экспериментах
пороговые концентрации белков выбирались значительно меньше ожидаемых
стационарных концентраций. В третьем эксперименте значения параметров
выбирались так же как и во втором, а пороговые концентрации белков были
значительно увеличены и по значениям "приближены" к ожидаемым стационарным
значениям концентраций. Для первого, второго и третьего экспериментов
компьютером было осуществлено 1000, 300 и 1000 случайных выборов
соответственно. Способность системы переходить к стационарному режиму и
находиться в нем оценивалась в конце выбранного отрезка времени
длительностью 96 часов по совокупности включенных генов в данном
стационарном состоянии, а также посредством анализа последовательности
включения генов во времени. Результаты вычислений приведены в таблице 1.
- параметр из набора
со значением, меньшим нижней границы соответствующего правдоподобного
интервала. Во втором эксперименте значения параметров случайно выбирались в
пределах 30% отклонения от "хорошего" набора параметров. В качестве
"хорошего" набора параметров выбран такой, в котором значения всех
переменных лежат в середине правдоподобных интервалов. В обоих экспериментах
пороговые концентрации белков выбирались значительно меньше ожидаемых
стационарных концентраций. В третьем эксперименте значения параметров
выбирались так же как и во втором, а пороговые концентрации белков были
значительно увеличены и по значениям "приближены" к ожидаемым стационарным
значениям концентраций. Для первого, второго и третьего экспериментов
компьютером было осуществлено 1000, 300 и 1000 случайных выборов
соответственно. Способность системы переходить к стационарному режиму и
находиться в нем оценивалась в конце выбранного отрезка времени
длительностью 96 часов по совокупности включенных генов в данном
стационарном состоянии, а также посредством анализа последовательности
включения генов во времени. Результаты вычислений приведены в таблице 1.
| № эксперимента | Нормальных режимов ( % ) в мутовках w |
Нормальных цветков (% ) |
|||
| чашелистики | лепестки | тычинки | плодолистики | ||
| 1 | 99.5 | 96.7 | 84.6 | 93.2 | 74 |
| 2 | 100 | 100 | 100 | 100 | 100 |
| 3 | 63.3 | 25.3 | 47 | 33.8 | - |
Интерпретация. (1) В условиях первого и второго экспериментов наблюдается высокий процент нормальных режимов, то есть модель обладает параметрической устойчивостью. Для AT-ПУМ существуют такие наборы кинетических параметров, что при одном и том же поведении на качественном уровне, система может обладать сильно различающимися количественными характеристиками (2-10-кратные различия в уровне концентраций молекулярных компонент). (2) В условиях третьего эксперимента модель не обладает параметрической устойчивостью. На этом основании можно предположить, что данное соотношение между коэффициентами синтеза продуктов генов и пороговыми концентрациями белков в изучаемой системе нереалистично. (3) Сравнение результатов всех трех экспериментов показывает, что с приближением значений порогов к значениям концентраций белков в стационарных состояниях устойчивость системы понижается.
Качественное описание системы. План строения тела Drosophila и дифференцированные структуры детерминируются уже на
стадии формирования клеточной бластодермы (Сингер, Берг, 1998).
Дифференциальная экспрессия генов на ранних стадиях эмбрионального развития
Drosophila в значительной мере связана с регуляцией транскрипции генов пяти классов -
maternal coordinate, gap, pair rule, segment polarity и homeotic. Гены
каждого класса находятся под воздействием генов, функционирующих на более
ранних стадиях, и некоторых генов своего класса, так что смена стадий
формирования тела Drosophila в процессе эмбриогенеза определяется иерархической
регуляцией по времени. Рассматриваемая подсеть состоит из 22 генов: bicoid
(bcd), caudal (cad), nanos (nos) класса maternal coordinate, giant (gt), hunchback (hb), knirps (kni), kruppel (kr), tailless (tll) класса gap,
even-skipped (eve), fushi tarazu (ftz), hairy (![]() , odd-skipped (odd), odd-paired (opa), paired (prd), runt (run) класса pair rule, engrailed (en), wingless (wg) класса segment
polarity, abdominal-A (abd-A), antennapedia (antp), deformed (dfd), sex combs reduced (scr), ultrabithorax (ubx) класса homeotic. Выбор данной совокупности
генов обусловлен тем, что каждый из них тесно вовлечен в процессы прямой и
опосредованной регуляции экспрессии генов, а также имеет сайты, специфичные
регуляторным молекулам белковых продуктов отдельных генов сети (Brody, 1999,
банк данных; Spirov et al., 2000, банк данных), и играет ключевую роль в
процессе формирования клеточной бластодермы и сегментации тела Drosophila.
, odd-skipped (odd), odd-paired (opa), paired (prd), runt (run) класса pair rule, engrailed (en), wingless (wg) класса segment
polarity, abdominal-A (abd-A), antennapedia (antp), deformed (dfd), sex combs reduced (scr), ultrabithorax (ubx) класса homeotic. Выбор данной совокупности
генов обусловлен тем, что каждый из них тесно вовлечен в процессы прямой и
опосредованной регуляции экспрессии генов, а также имеет сайты, специфичные
регуляторным молекулам белковых продуктов отдельных генов сети (Brody, 1999,
банк данных; Spirov et al., 2000, банк данных), и играет ключевую роль в
процессе формирования клеточной бластодермы и сегментации тела Drosophila.
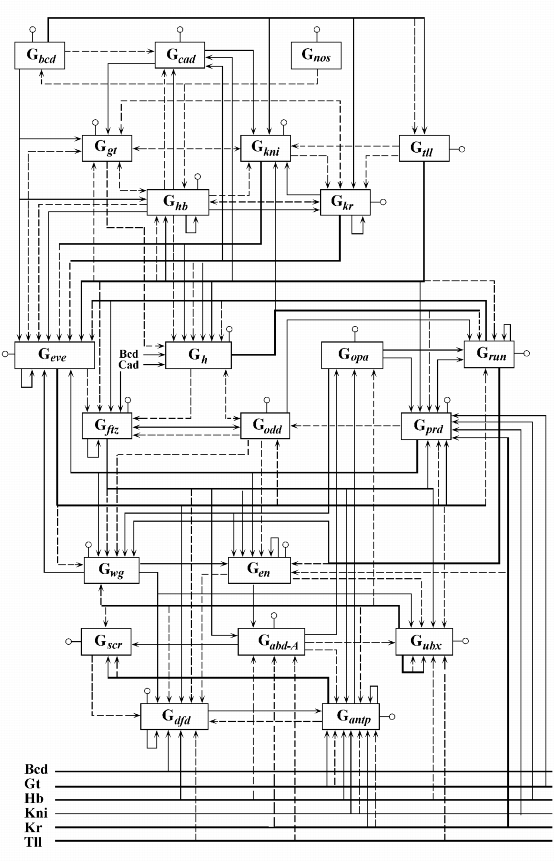
Анализ модели. В модель Dr-УГС-1 входят синтетические блоки G![]() , G
, G![]() ,
..., G
,
..., G
![]() - блоки синтеза белковых продуктов генов
bcd, nos, gt, hb, kni, kr, tll, eve, ftz,
- блоки синтеза белковых продуктов генов
bcd, nos, gt, hb, kni, kr, tll, eve, ftz, ![]() , odd, opa, prd, run, en, wg, abd-A, antp, dfd, scr, ubx, cad соответственно. Блок-схема модели
изображена на рис. 4.
, odd, opa, prd, run, en, wg, abd-A, antp, dfd, scr, ubx, cad соответственно. Блок-схема модели
изображена на рис. 4.
 |
Также как и для модели AT-ПУМ-2 посредством компьютерных расчетов на модели Dr-УГС-1 были выявлены стационарные состояния изучаемой генной сети, получены кинетические кривые для белков, описывающие динамику системы при переходах в стационарные состояния. На рис. 5 представлены кинетические кривые для белков, отражающие динамику фрагмента генной сети, состоящего из генов класса maternal и gap, в четвертом парасегменте яйца Drosophila.
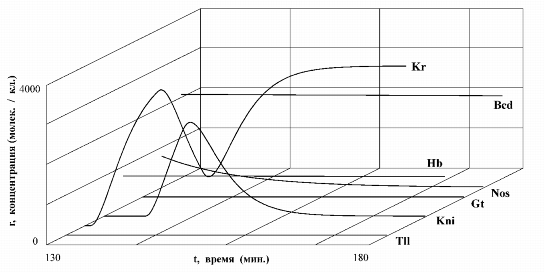 |
Каждому стационарному состоянию
![]() модели
Dr-УГС-1 с соответствующими наборами кинетических кривых и диаграмм
активностей генетических блоков можно сопоставить однозначные паттерны
экспрессии генов в определенной части яйца, которые формируется в процессе
реально протекающего раннего онтогенеза Drosophila (рис. 6). Таким образом, кинетика
модели Dr-УГС-1 "спроецирована" на пространственные компартменты эмбриона
Drosophila.
модели
Dr-УГС-1 с соответствующими наборами кинетических кривых и диаграмм
активностей генетических блоков можно сопоставить однозначные паттерны
экспрессии генов в определенной части яйца, которые формируется в процессе
реально протекающего раннего онтогенеза Drosophila (рис. 6). Таким образом, кинетика
модели Dr-УГС-1 "спроецирована" на пространственные компартменты эмбриона
Drosophila.
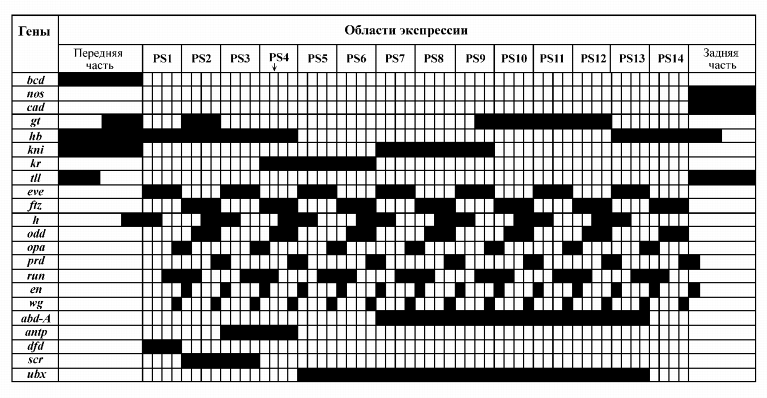 |
Каждая вертикальная полоска на рисунке в области 1PS - 14PS (PS - парасегмент) соответствует одному ядру бластодермы. В ядре, отмеченном стрелкой (область PS4), фрагмент генной сети, состоящий из генов классов maternal и gap, имеет динамику, представленную на рис. 5.
Использованные при построении моделей AT-ПУМ-2 и Dr-УГС-1 принципы допускают расширение моделей с учетом дополнительных информационных связей, генов других классов, а для Dr-УГС-1 - также ооплазматической биохимической сегрегации, ярко представленной в фертилизированном яйце Drosophila. Модели обладают прогностической ценностью, поскольку позволяют выявлять режимы поведения генных сетей, находить стационарные состояния, проверять их параметрическую устойчивость, отслеживать траектории изменения количества молекулярных компонент сети, изучать особенности переходов из одного функционального состояния сети в другое, предсказывать вид кинетических кривых для генных продуктов, а так же дискриминировать гипотезы о возможных механизмах взаимодействия между генами и регуляторными веществами.
 Ваши комментарии
Ваши комментарии
|
![[SBRAS]](/gif/s_sigma.gif)
[Головная страница] [Конференции] [СО РАН] |
© 2001, Сибирское отделение Российской академии наук, Новосибирск
© 2001, Объединенный институт информатики СО РАН, Новосибирск
© 2001, Институт вычислительных технологий СО РАН, Новосибирск
© 2001, Институт систем информатики СО РАН, Новосибирск
© 2001, Институт математики СО РАН, Новосибирск
© 2001, Институт цитологии и генетики СО РАН, Новосибирск
© 2001, Институт вычислительной математики и математической геофизики СО РАН, Новосибирск
© 2001, Новосибирский государственный университет
Дата последней модификации Tuesday, 11-Sep-2001 14:57:35 NOVST