
Хидиров Б.Н.
Институт кибернетики, Ташкент
One of the possible approaches for quantitative studies of
regulatory mechanisms of living system is considered. The equations of
system state are constructed taking into account the cooperation of
processes, presence of combined feedback and temporary relations in
regulation loop. Results of the qualitative analysis of an equations
and their model systems shows existence of following functioning
regimes: rest, stable functional state, periodic and irregular
fluctuations. In some cases the effect "black hole'' - break-down of
solutions to trivial attractor is observed. The results application of
model investigations for the analysis some biological problems shows,
according to our opinion, that the proposed method can be used for
quantitative studies of living system mechanisms at norm and
anomalies.
Математические модели живых систем предназначены для количественного
анализа поведения множества элементов (комплекса макромолекул),
функционирующих в некоторой особой среде и способных реагировать на
определенные внешние воздействия. Это приводит [18,19] к
формулировке
понятий ![]() (операторов-регуляторов) - элементов регуляторной системы,
способной к восприятию и выработке сигналов определенной природы, и
(операторов-регуляторов) - элементов регуляторной системы,
способной к восприятию и выработке сигналов определенной природы, и
![]() (active system with time average) - сигнальной среды регуляторной
системы, в которой взаимосвязанная деятельность элементов
осуществляется, на основе обратных связей, с некоторым средним
временем h (временем, прошедшим с момента образования сигналов до
момента воздействия их (или их продуктов) на активность элементов)
[18]. Комплекс
(active system with time average) - сигнальной среды регуляторной
системы, в которой взаимосвязанная деятельность элементов
осуществляется, на основе обратных связей, с некоторым средним
временем h (временем, прошедшим с момента образования сигналов до
момента воздействия их (или их продуктов) на активность элементов)
[18]. Комплекс ![]() вместе с
вместе с ![]() составляет регуляторную систему -
составляет регуляторную систему -
![]() . Геометрия подобных регуляторных систем является динамической,
в которой понятие неподвижной точки теряет смысл [19]. Единицей
времени
. Геометрия подобных регуляторных систем является динамической,
в которой понятие неподвижной точки теряет смысл [19]. Единицей
времени ![]() является
является ![]() . В природе наблюдаются случаи раздельного
существования
. В природе наблюдаются случаи раздельного
существования ![]() и
и ![]() . Примером
. Примером ![]() без
без ![]() являются вирусы,
а
являются вирусы,
а ![]() без
без ![]() - зрелые безъядерные эритроциты.
- зрелые безъядерные эритроциты.
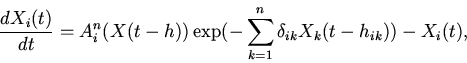 |
(1) |
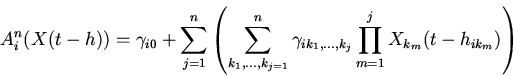
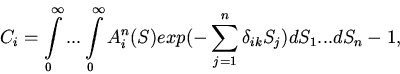 |
(2) |
Уравнение (1) относится к классу функционально-дифференциальных уравнений и,
при задании непрерывных функций на начальном временном отрезке длины
![]() ,
ее непрерывное решение может быть получено методом последовательного
интегрирования [16]. При реализации данного метода на современных ЭВМ
возникает задача построения решений по заданным дискретным значениям
искомых переменных, которая является актуальной и при количественном
описании биологических процессов функционально-дифференциальными
уравнениями при наличии только дискретных экспериментальных данных.
Данная задача для определенного класса линейных функционально-
дифференциальных уравнений является разрешимой и разработанный метод
построения их решений [16] можно использовать при количественном анализе
поведения решений (1).
,
ее непрерывное решение может быть получено методом последовательного
интегрирования [16]. При реализации данного метода на современных ЭВМ
возникает задача построения решений по заданным дискретным значениям
искомых переменных, которая является актуальной и при количественном
описании биологических процессов функционально-дифференциальными
уравнениями при наличии только дискретных экспериментальных данных.
Данная задача для определенного класса линейных функционально-
дифференциальных уравнений является разрешимой и разработанный метод
построения их решений [16] можно использовать при количественном анализе
поведения решений (1).
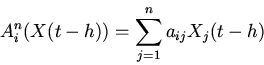
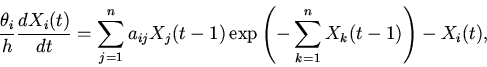 |
(3) |
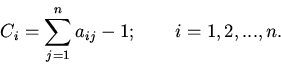
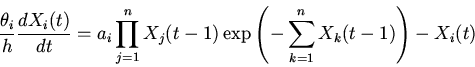 |
(4) |
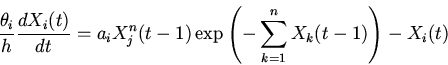 |
(5) |
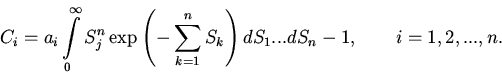
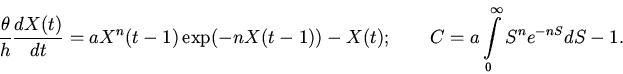 |
(6) |
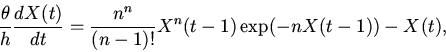 |
(7) |
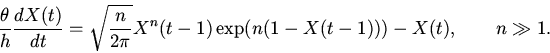 |
(8) |
Анализ характера решений уравнений (8), на основе методов качественного исследования функционально-дифференциальных уравнений [16,18,19], показывает нахождение их решений в первом квадранте фазового пространства при неотрицательности значений параметров и начальных условий; неустойчивость бесконечно удаленных точек; возможность существования тривиального и нетривиальных положений равновесия. Поведение решений в области нетривиального положения равновесия, являющегося функционально активным состоянием регуляторной системы, может носить характер устойчивых стационарных режимов, предельных циклов типа Пуанкаре и сложных колебаний [14].
Рассмотрим некоторые основные закономерности функционирования регуляторных механизмов (регуляторики) самосопряженных систем на основе качественного анализа уравнений (8).
Если примем, что, в случае нормальной регуляторики, сигналы,
вырабатываемые ![]() и в среднем существующие
и в среднем существующие ![]() единиц времени, должны
"дожить'' до осуществления петли обратной связи (т.е.
единиц времени, должны
"дожить'' до осуществления петли обратной связи (т.е. ![]() ),
то для них (8) примет вид
),
то для них (8) примет вид
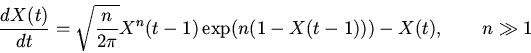 |
(9) |
Случай ![]() , когда сигнал (продукт активности
, когда сигнал (продукт активности ![]() )
может участвовать в
регуляции
)
может участвовать в
регуляции ![]() более чем один раз (т.к. сигнал живет дольше по сравнению
со временем, необходимым для осуществления петли
обратной связи), мы можем назвать сильной регуляторикой. Тогда
естественно назвать слабой регуляторикой случай
более чем один раз (т.к. сигнал живет дольше по сравнению
со временем, необходимым для осуществления петли
обратной связи), мы можем назвать сильной регуляторикой. Тогда
естественно назвать слабой регуляторикой случай ![]() . При очень слабой
регуляторике (
. При очень слабой
регуляторике (![]() ) для предварительного качественного анализа
рассматриваемых регуляторных систем может быть полезно следующее
функциональное уравнение регуляторики самосопряженных систем
[18,19]
) для предварительного качественного анализа
рассматриваемых регуляторных систем может быть полезно следующее
функциональное уравнение регуляторики самосопряженных систем
[18,19]
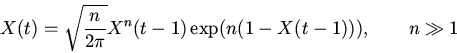 |
(10) |
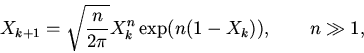 |
(11) |
Условие существования стационарных режимов функционирования
нетривиальных (положений равновесия в первом квадранте фазового пространства)
для рассматриваемых систем выражается в виде
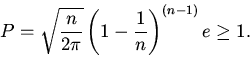 |
(12) |
При ![]() имеется только тривиальное положение равновесия. Нетривиальное
положение равновесия возникает, путем жесткого возбуждения, при
имеется только тривиальное положение равновесия. Нетривиальное
положение равновесия возникает, путем жесткого возбуждения, при ![]() в
точке
в
точке ![]() . Рост значения
. Рост значения ![]() приводит к бифуркации положения
равновесия
приводит к бифуркации положения
равновесия ![]() на два нетривиальных положений равновесия
на два нетривиальных положений равновесия ![]() , постепенно
удаляющихся от точки
, постепенно
удаляющихся от точки ![]() на две стороны. Расчеты на ЭВМ показывают (с
учетом принятого приближения, на основе формулы Стирлинга для больших
на две стороны. Расчеты на ЭВМ показывают (с
учетом принятого приближения, на основе формулы Стирлинга для больших
![]() [19]), что нетривиальные положения равновесия самосопряженных
регуляториных систем возникают при
[19]), что нетривиальные положения равновесия самосопряженных
регуляториных систем возникают при ![]() и, по мере роста
и, по мере роста ![]() , вначале
удаляются от точки
, вначале
удаляются от точки ![]() , затем сближаются друг с другом, оставаясь в разных
сторонах от
, затем сближаются друг с другом, оставаясь в разных
сторонах от ![]() .
.
Рассмотрим устойчивость положений равновесия при ![]() . Очевидна
устойчивость тривиального положения равновесия. Использование метода
Ляпунова, путем линеаризации (8) около нетривиального положения
равновесия
. Очевидна
устойчивость тривиального положения равновесия. Использование метода
Ляпунова, путем линеаризации (8) около нетривиального положения
равновесия ![]() , приводит к характеристическому уравнению
, приводит к характеристическому уравнению
| (13) |
Для случая систем с очень слабой регуляторикой получаем условие
устойчивости ![]() в виде неравенства
в виде неравенства ![]() . Как показывают расчеты,
уже для
. Как показывают расчеты,
уже для ![]() оно не выполняется, т.е. при
оно не выполняется, т.е. при ![]() устойчивость
положения
равновесия в точке
устойчивость
положения
равновесия в точке ![]() нарушается. Результаты качественного исследования
решений (11) в области неустойчивости показывают существование
довольно сложного диапазона поведения рассматриваемой модели
самосопряженных регуляторных систем. Помимо периодических режимов
функционирования, возможны нерегулярные колебания и эффект "черная
дыра'' [19]. Последний заключается в возникновении деструктивных
изменений в моделируемой системе и выражается срывом колебаний -
стремлением решений к тривиальному положению равновесия. Расчеты,
проведенные на ПЭВМ с помощью вычисления Ляпуновской величины,
информационоой и высших размерностей решений, построения диаграмм
Кенингса-Ламерея (11), показывают существование нерегулярных колебаний
при
нарушается. Результаты качественного исследования
решений (11) в области неустойчивости показывают существование
довольно сложного диапазона поведения рассматриваемой модели
самосопряженных регуляторных систем. Помимо периодических режимов
функционирования, возможны нерегулярные колебания и эффект "черная
дыра'' [19]. Последний заключается в возникновении деструктивных
изменений в моделируемой системе и выражается срывом колебаний -
стремлением решений к тривиальному положению равновесия. Расчеты,
проведенные на ПЭВМ с помощью вычисления Ляпуновской величины,
информационоой и высших размерностей решений, построения диаграмм
Кенингса-Ламерея (11), показывают существование нерегулярных колебаний
при ![]() и наступление эффекта "черная дыра'' при
и наступление эффекта "черная дыра'' при ![]() . Следовательно,
стабильные самосопряженные регуляторные системы с очень слабой
регуляторикой, находящиеся в равновесии с внешней средой, могут
существовать только при
. Следовательно,
стабильные самосопряженные регуляторные системы с очень слабой
регуляторикой, находящиеся в равновесии с внешней средой, могут
существовать только при
![]() [18,19].
[18,19].
В первом случае рассматривались закономерности роста клеток
хлореллы в связи с задачей оптимизации их выращивания [14]. Применение
рассматриваемого подхода показало, что онтогенез хлореллы (рост молодой
клетки (до материнской) ![]() деление-дробление
деление-дробление ![]() выход молодых клеток
хлореллы во внешнюю среду) сопровождается увеличением
самосопряженности популяции клеток при размножении. Достижение
самосопряженности (количества клеток) порогового значения (12) приводит к
эффекту "черная дыра'', т.е. к распаду системы, что и наблюдается при
экспериментах после 4- 5-ти кратного деления [14].
выход молодых клеток
хлореллы во внешнюю среду) сопровождается увеличением
самосопряженности популяции клеток при размножении. Достижение
самосопряженности (количества клеток) порогового значения (12) приводит к
эффекту "черная дыра'', т.е. к распаду системы, что и наблюдается при
экспериментах после 4- 5-ти кратного деления [14].
Во втором случае был проведен анализ механизмов возникновения
злокачественного новообразования. Была рассмотрена гипотеза об управлени
ранним эмбриональным развитием посредством продуктов (и-РНК)
генетической Системы Автономного Развития (САР). САР функционирует
на этапе "ламповых щеток'' оогенеза для образования необходимого
количества соответствующих и-РНК (и хранящихся в виде информасом).
Они используются, после оплодотворения, для обеспечения развития
зародыша автономно от материнского организма до включения
собственного генома (обычно до гаструлы [17]). Как показали результаты
модельных исследований на основе рассматриваемых уравнений
регуляторики живых систем [20], САР может состоять из нескольких блоков
(инициации, образования и-РНК и репрессии) и жестко блокируется после
оогенеза. Деблокировка САР, на основе внутренних и внешних причин,
приводит к появлению последовательных залпов генетической информации
САР и развитию автономных клеток, которые могут дать начало
злокачественному росту [17,20]. Продукты блока инициации,
возникающие до
автономного развития, могут быть использованы для раннего обнаружения
активации САР, а продукты блока репрессии - для подавления
функциональной САР [17,20].
В условиях равновесия с внешней средой самосопряженная регуляторная система, с очень слабой регуляторикой, обладает семью вариантами стабильного существования, когда степень самосопряжения принимает значения от шести до двенадцати. Более высокие сопряженности могут существовать при организации иерархических регуляторных систем (что и наблюдается в реальных условиях). Использование данного подхода модельного исследования регуляторных систем для анализа механизмов развития микроорганизмов [14,15] и ракового новообразования [1,20] показало его приемлемость при количественном исследовании механизмов регуляции биосистем.
 Ваши комментарии
Ваши комментарии
|
![[SBRAS]](/gif/s_sigma.gif)
[Головная страница] [Конференции] [СО РАН] |
© 2001, Сибирское отделение Российской академии наук, Новосибирск
© 2001, Объединенный институт информатики СО РАН, Новосибирск
© 2001, Институт вычислительных технологий СО РАН, Новосибирск
© 2001, Институт систем информатики СО РАН, Новосибирск
© 2001, Институт математики СО РАН, Новосибирск
© 2001, Институт цитологии и генетики СО РАН, Новосибирск
© 2001, Институт вычислительной математики и математической геофизики СО РАН, Новосибирск
© 2001, Новосибирский государственный университет
Дата последней модификации Tuesday, 11-Sep-2001 19:18:44 NOVST