| (1) |

Бандман О.Л.
Институт вычислительной математики
и математической геофизики СО РАН, Новосибирск
A fine-grained parallel algorithm is proposed, which is intended for
simulation spatial dynamics in active media. The algorithm
combines into a single iterative procedure Boolean operations
of Cellular Automaton Diffusion with Cellular-Neural
simulation of nonlinear reaction of active medium.
Methods for transforming a continuous spatial function into its
boolean equivalent (Boolean discretization), and the inverse methods
to transform a Boolean space into a function in real numbers
(averaging) are elaborated to be used on each iteration.
Such a "hybrid'' (discrete + continuous) algorithm is absolutely
stable relative to round off errors and allows natural
parallelization. Examples are given to show simulation of nonlinear
waves in ecological system.
Процессы динамики в активных средах (реакционно-диффузионные процессы в физике, химии, биологии, экологии [6,7]) традиционно представляются дифференциальными уравнениями (или системами уравнений) с частными производными. Правая часть этих уравнений состоит из двух частей: диффузионной, представленной лапласианом, и реакционной, представленной нелинейной функцией, отображающей активную составляющую. Уравнения этого типа изучаются "поштучно'' и с большим трудом, так как аналитические решения получить невозможно из-за нелинейного члена, а численные методы либо плохо распараллеливаются (неявные схемы решений), либо ограничены условиями устойчивости и точности (явные схемы). А между тем алгоритмы КА-диффузии таких ограничений не имеют. Они достаточно хорошо разработаны и теоретически обоснованы [8,2]. Отсюда вытекает стремление использовать дискретные КА- алгоритмы для моделирования диффузионной составляющей и клеточно-нейронную (КН) модель [5] - для нелинейной реакции. В статье предлагается такая "гибридная'' модель и делается попытка ее обосновать теоретически и подтвердить экспериментально.
Кроме введения, статья содержит 4 раздела и заключение. Во втором разделе дана формальная постановка задачи и общая схема предлагаемого алгоритма. В третьем разделе представлены методы КА-диффузии и КН-реакции. В четвертом - приведены результаты экспериментов. В заключении намечены проблемы для дальнейших исследований.
| (1) |
| (2) |
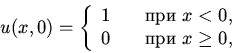 |
(3) |
| (4) |
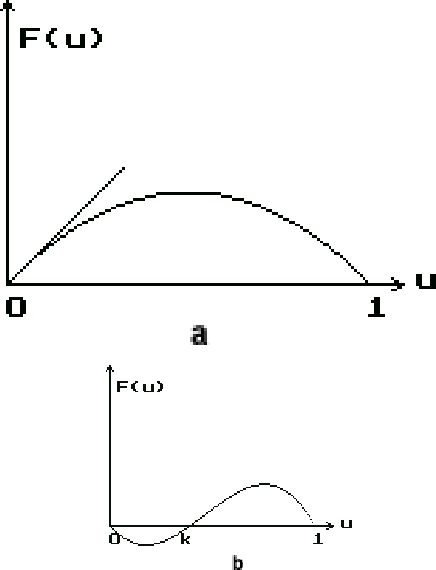
В исследованиях по экологии функции, удовлетворяющие условиям (2),
называются логистическими и считаются базовыми, хотя
применяются также полиномы третьей степени (рис.1b)
| (5) |
При начальных условиях (3) и ![]() вида (5)
скорость бегущего фронта
вида (5)
скорость бегущего фронта
| (6) |
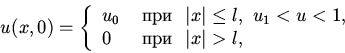 |
(7) |
Уравнения второго порядка моделируют более сложные явления
(химические реакции, распространение волн, эпидемий,
образование вихрей и др.) Например, модель Тайсона-Файфа реакции
Белоусова--Жаботинского [6], порождающей концентрические
расходящиеся волны.
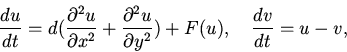 |
(8) |
Представленные выше примеры реакционно-диффузионных уравнений и их известных свойств затем будут использованы для сравнения с аналогичными свойствами, полученными с помощью модели клеточно-нейронного автомата.
Как и для конечно-разностных методов решения дифференциальных
уравнений с частными производными, КН-модель содержит процедуры
дискретизации пространственных координат и времени,
так что
![]() . При
. При ![]() уравнение (1)
приобретает вид
уравнение (1)
приобретает вид
| (9) |
| (10) |
Клеточно-нейронная модель (также как и автоматная)
имеют дело с
дискретными величинами основной
переменной, которые являются "предельно дискретными'', т.е. булевыми.
Поскольку КН-автомат использует и дискретные и непрерывные переменные, то для описания его работы далее применяются
Формализмы универсальной модели клеточных
Вычислений - алгоритма параллельных подстановок (АПП) [1]. В этой
модели функция ![]() представляется в
виде клеточного множества
представляется в
виде клеточного множества ![]() , элементами которого
служат пары вида
, элементами которого
служат пары вида ![]() , называемые клетками. В них
, называемые клетками. В них ![]() A - символ алфавита A, обозначающий состояние клетки, а
A - символ алфавита A, обозначающий состояние клетки, а
![]() M - имя клетки из множества имен M. Множество
имен (в общем случае, счетное), обычно, состоит из номеров клеток
или наборов пространственных координат. Например, если функция
M - имя клетки из множества имен M. Множество
имен (в общем случае, счетное), обычно, состоит из номеров клеток
или наборов пространственных координат. Например, если функция
![]() определена в двумерном декартовом пространстве
определена в двумерном декартовом пространстве
![]() , то множество имен M
, то множество имен M
![]() . В каждой точке с именем
. В каждой точке с именем ![]() значение функции
значение функции ![]() является состоянием клетки
является состоянием клетки ![]() .
.
Операции над клеточными множествами задаются системами
параллельных подстановок вида
| (11) |
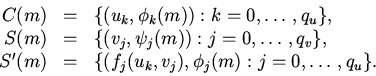 |
(12) |
Параллельные подстановки применяются все одновременно и
параллельно ко всем тем клеткам клеточного множества, для которых
локальная конфигурация совпадает с левой частью подстановки. В
результате их применения состояния клеток базовой части ![]() заменяются на результат вычисления функций
заменяются на результат вычисления функций ![]() .
Клетки локальной конфигурации
.
Клетки локальной конфигурации ![]() , называемой контекстом
не меняют состояний. Такое итерационное применение подстановок
моделирует эволюцию клеточного множества и продолжается до тех пор,
пока система не придет в устойчивое состояние.
, называемой контекстом
не меняют состояний. Такое итерационное применение подстановок
моделирует эволюцию клеточного множества и продолжается до тех пор,
пока система не придет в устойчивое состояние.
Исходное состояние может быть задано
клеточным множеством
![]() , где
, где ![]() A - булевы константы.
или вещественной функцией
A - булевы константы.
или вещественной функцией ![]() в дискретизированном пространстве.
В первом случае чтобы получить вещественные значения необходимо
выполнить над
в дискретизированном пространстве.
В первом случае чтобы получить вещественные значения необходимо
выполнить над ![]() подстановку
подстановку
![]() , где
, где
![]() R,
в котором значения
R,
в котором значения ![]() получены путем применения
процедуры
осреднения,
получены путем применения
процедуры
осреднения,
![]()
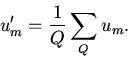 |
(13) |
Функционирование
КН-автомата моделирует итерационнный
алгоритм конечно-разностного уравнения (10),
на каждом шаге вычисляя первые два члена правой части как
КА-диффузию, и третий член как
''клеточно-нейронную реакцию''.
Формально работа КН-автомата описывается следующим образом.
Пусть результат ![]() -го шага в булевом
представлении равен клеточному множеству
-го шага в булевом
представлении равен клеточному множеству
![]() . Один итерационный шаг КН-автомата
состоит из следующих двух операций.
. Один итерационный шаг КН-автомата
состоит из следующих двух операций.
1. Выполнение одного шага КА-диффузии (раздел 3.1).
2. Выполнение одного шага клеточно-нейронной реакции.
При этом клетка выполняет функцию
вероятностного нейрона (раздел 3.2), которая состоит в
том, чтобы изменить булево состояние клетки в зависимости от значения
![]() .
.
Исходным является клеточное множество
![]() ,
которое может быть получено путем применения процедуры булевой
дискретизации к заданной пространственной функции
распределения концентрации. Строится два разбиения множества имен
M{[i,j]}на
блоки, размером 2х2: четное разбиение, в котором диагональные
клетки блоков имеют четную сумму координат (
,
которое может быть получено путем применения процедуры булевой
дискретизации к заданной пространственной функции
распределения концентрации. Строится два разбиения множества имен
M{[i,j]}на
блоки, размером 2х2: четное разбиение, в котором диагональные
клетки блоков имеют четную сумму координат (![]() ), и
нечетное, в котором диагональные клетки блоков имеют нечетную
сумму координат (
), и
нечетное, в котором диагональные клетки блоков имеют нечетную
сумму координат (![]() ). Каждая итерация алгоритма разделена
на два шага. На четных шагах клеточно-автоматное правило перехода
применяется к четным блокам, на нечетных - то же самое правило
применяется к нечетным. Правило состоит в следующем. На каждом четном (нечетном)
шаге все четные (нечетные) блоки поворачиваются на
). Каждая итерация алгоритма разделена
на два шага. На четных шагах клеточно-автоматное правило перехода
применяется к четным блокам, на нечетных - то же самое правило
применяется к нечетным. Правило состоит в следующем. На каждом четном (нечетном)
шаге все четные (нечетные) блоки поворачиваются на ![]() по часовой или против часовой стрелки с равной вероятностью. В
терминах АПП это выражается формально в виде системы
из двух подстановок
по часовой или против часовой стрелки с равной вероятностью. В
терминах АПП это выражается формально в виде системы
из двух подстановок
![]() , где
, где
![\begin{displaymath}
\left \{ \begin{array}{lll}
\Theta_1:& \{(rand(1)<p,[i,j]...
...), (u_3,[i+1,j+1]),
(u_0,[i+1,j]), \\
\end{array} \right.
\end{displaymath}](img73.gif) |
(14) |
Одномерный ПБ-метод отличается от двумерного тем, что блоки в нем состоят из пар клеток, причем четные (нечетные) блоки имеют четное (нечетное) имя левой клетки. На четных (нечетных) шагах клетки четных (нечетных) блоков обмениваются состояниями.
| (15) |
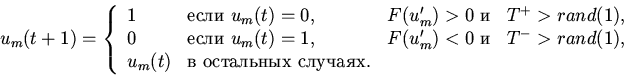 |
(16) |
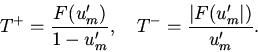 |
(17) |
| (18) |
Точного решения поставленная задача не имеет. Приближенно она
решается следующим образом. Состояние каждой клетки
определяется из условия: вероятность того, что оно равно 1
равна ![]()
| (19) |
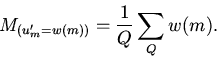 |
(20) |
Предположим, что функция в области соседства клетки постоянна, т.е.
![]() .
Тогда, согласно (20) и (21)
.
Тогда, согласно (20) и (21)
![]() .
Следовательно, в этом вырожденном случае ошибка булевой дискретизации
.
Следовательно, в этом вырожденном случае ошибка булевой дискретизации
![]() . Пусть
среднее отклонение состояний клеток в окрестности осреднения
. Пусть
среднее отклонение состояний клеток в окрестности осреднения
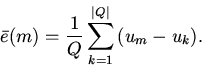 |
(21) |
| (22) |
Из (22) можно сделать заключение, что если функция имеет постоянную
пространственную производную в окрестности осреднения точки ![]() (функция
(функция ![]() линейна), то
линейна), то ![]() , так
как положительные и отрицательные отклонения компенсируют друг
друга. По той же причине
, так
как положительные и отрицательные отклонения компенсируют друг
друга. По той же причине ![]() для функций вида
для функций вида
![]() , (
, (![]() - произвольные константы) и
всех остальных парабол нечетной степени.
- произвольные константы) и
всех остальных парабол нечетной степени.
Точность булевой дискретизации зависит от выбора двух параметров:
1) размеров пространственного шага (будем считать что он
одинаков по всем координатным направлениям и равен ![]() ),
),
2) мощности окрестности осреднения ![]() , которая в случае
1D равна
, которая в случае
1D равна ![]() , а в случае 2D равеа
, а в случае 2D равеа ![]() (
(![]() -радиус осреднения).
-радиус осреднения).
Эти параметры должны быть выбраны так, чтобы, с одной стороны, минимизировать "автоматный шум'', и, с другой стороны, чтобы была минимальной ошибка осреднения. Эти два требования противоречивы, первое требует большой окрестности, второе - маленькой.
Требование малого автоматного шума может быть выражено следующим
образом:
| (23) |
Требование малой ошибки осреднения формулируется исходя из
того, что на каждом пространственном шаге отклонение от
от средней ошибки равно
| (24) |
| (25) |
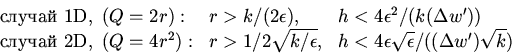 |
(26) |
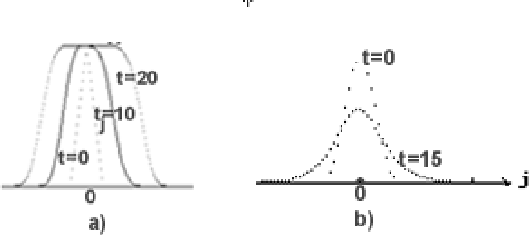
Моделировался процесс распространения концентрации, описанный
типовым уравнением (1), c реакционной
составляющей вида (2) при ![]() ,
, ![]() и начальными
условиями типа "вспышки''. Размеры клеточного множества 32х256
клеток. Распространение фронта происходит вдоль оси
и начальными
условиями типа "вспышки''. Размеры клеточного множества 32х256
клеток. Распространение фронта происходит вдоль оси ![]() ,
,
![]() . В каждом подмножестве клеток с
. В каждом подмножестве клеток с
![]() , (строке клеточной структуры) выполняется алгоритм
одномерной КА-диффузии. Осреднение (вычисление
, (строке клеточной структуры) выполняется алгоритм
одномерной КА-диффузии. Осреднение (вычисление ![]() ) по
окрестности
) по
окрестности
![]() . Функции КН-реакции
вычисляется по алгоритму вероятностного нейрона (16) с вероятностями
. Функции КН-реакции
вычисляется по алгоритму вероятностного нейрона (16) с вероятностями
![]() . По расстояниями между фронтами при разных
. По расстояниями между фронтами при разных ![]() определялась скорость распространения, которая постепенно
увеличивалась с увеличением времени,
начиная с
определялась скорость распространения, которая постепенно
увеличивалась с увеличением времени,
начиная с ![]() при
при ![]() и кончая
и кончая ![]() при
при ![]() ,
(рис.2.a) (для сравнения по формуле (4)
,
(рис.2.a) (для сравнения по формуле (4)
![]() .
Форма фронта совпадает с расчитанной по (4) с точностью до
.
Форма фронта совпадает с расчитанной по (4) с точностью до ![]() .
То же уравнение c теми же начальными условиями и параметрами
моделирования, но с
.
То же уравнение c теми же начальными условиями и параметрами
моделирования, но с ![]() вида (5) дали затухающую вспышку
(рис.2b).
вида (5) дали затухающую вспышку
(рис.2b).
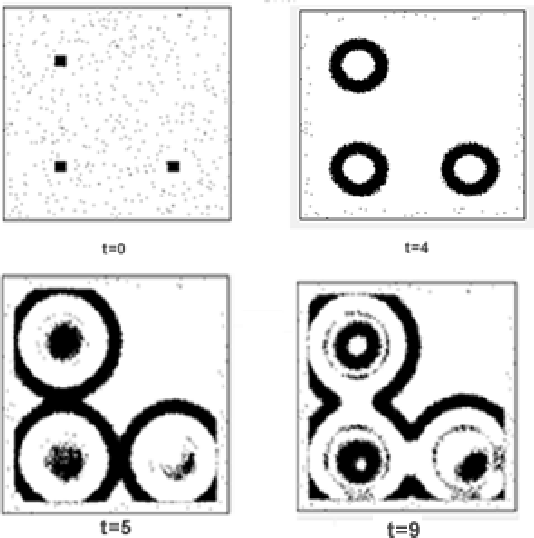
Моделировалась система уравнений (8) на клеточном множестве
размерами 200х200 при ![]() и начальных концентрациях
и начальных концентрациях
![]() на всем моделируемом пространстве, кроме трех точек
возбуждения (рис.3). КА-диффузия выполнялась двумерным алгоритмом
поворота блоков, КН-реакция - по формулам (17). На рис. 3 показаны
возникающие вокруг точек возбуждения концентрические волны.
на всем моделируемом пространстве, кроме трех точек
возбуждения (рис.3). КА-диффузия выполнялась двумерным алгоритмом
поворота блоков, КН-реакция - по формулам (17). На рис. 3 показаны
возникающие вокруг точек возбуждения концентрические волны.
 Ваши комментарии
Ваши комментарии
|
![[SBRAS]](/gif/s_sigma.gif)
[Головная страница] [Конференции] [СО РАН] |
© 2001, Сибирское отделение Российской академии наук, Новосибирск
© 2001, Объединенный институт информатики СО РАН, Новосибирск
© 2001, Институт вычислительных технологий СО РАН, Новосибирск
© 2001, Институт систем информатики СО РАН, Новосибирск
© 2001, Институт математики СО РАН, Новосибирск
© 2001, Институт цитологии и генетики СО РАН, Новосибирск
© 2001, Институт вычислительной математики и математической геофизики СО РАН, Новосибирск
© 2001, Новосибирский государственный университет
Дата последней модификации Tuesday, 11-Sep-2001 16:54:34 NOVST